


Curr. Oncol. 2024, 31(5), 2488-2496; https://doi.org/10.3390/curroncol31050186 (registering DOI) - 28 Apr 2024
Abstract
Background: Unsolicited patient feedback (compliments and complaints) should allow the healthcare system to address and improve individual and overall patient, family, and staff experiences. We evaluated feedback at a tertiary cancer centre to identify potential areas for optimizing care delivery. Methods: unsolicited feedback
[...] Read more.
Background: Unsolicited patient feedback (compliments and complaints) should allow the healthcare system to address and improve individual and overall patient, family, and staff experiences. We evaluated feedback at a tertiary cancer centre to identify potential areas for optimizing care delivery. Methods: unsolicited feedback submitted to the Patient Relations Department, relating to the Divisions of Medical and Radiation Oncology, at the Ottawa Hospital, was analyzed. Results: Of 580 individual reports submitted from 2016 to 2022, patient demographics were available for 97% (563/580). Median patient age was 65 years (range 17–101), and 53% (301/563) were female. The most common cancer types were breast (127/545, 23%) and gastrointestinal (119/545, 22%) malignancies, and most (64%, 311/486) patients had metastatic disease. Feedback was submitted mainly by patients (291/579, 50%), and predominantly negative (489/569, 86%). The main reasons for complaints included: communication (29%, 162/566) and attitude/conduct of care (28%, 159/566). While feedback rates were initially stable, an increase occurred from 2019 to 2021. Conclusions: Unsolicited feedback remains mostly negative, and relates to physician communication. If we are to drive meaningful changes in care delivery, more standardized means of assessing feedback and implementation strategies are needed. In addition, in an era of increased healthcare provider burnout, strategies to enhance formal positive feedback are also warranted.
Full article
(This article belongs to the Special Issue The 30th Anniversary of Current Oncology: Perspectives in Clinical Oncology Practice)
►
Show Figures
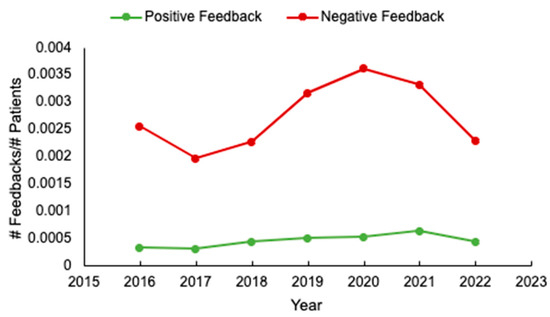
Figure 1





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}